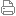
纹状体[5](也称为新纹状体和纹状核)是前脑皮质下基底神经节中的细胞核(神经元簇)。纹状体是运动和奖励系统的关键组成部分;接受来自不同来源的谷氨酸能和多巴胺能输入;并作为其他基底神经节的主要输入。
在功能上,纹状体协调认知的多个方面,包括运动和行动计划,决策,动机,强化和奖励感知。[2] [3] [4]纹状体由尾状核和豆状核组成。[6] [7]豆状核由较大的壳核和较小的苍白球构成。[8]
在灵长类动物中,纹状体分为腹侧纹状体和背侧纹状体,基于功能和连接的细分。腹侧纹状体由伏隔核和嗅结节组成。背侧纹状体由尾状核和壳核组成。背侧纹状体中的白质,神经束(内囊)将尾状核和壳核分开。[4]在解剖学上,术语纹状体描述了灰色和白色物质的条纹(条纹)外观。[9]
https://cache.tv.qq.com/qqplayerout.swf?vid=z0864tnrouj
视频:2分钟神经科学 - 纹状体

紫色=尾状和壳核,橙色=丘脑

显示皮质纹状体连接的纤维束成像
目录
1 结构
1.1 细胞类型
1.2 输入
1.3 目标
2 功能
3 临床意义
3.1 帕金森病
3.2 成瘾
3.3 双相情感障碍
3.4 自闭症谱系障碍
3.5 功能障碍
4 历史
5 其他动物
6 参考
结构
在MRI上看到的纹状体。 纹状体包括尾状核和豆状核,其中包括壳核和苍白球。

在MRI上看到的红色纹状体。纹状体包括尾状核(顶部)和豆状核(壳核(右)和左下苍白球)
纹状体是基底神经节的最大结构。根据功能和连接,纹状体分为腹侧和背侧分区。
腹侧纹状体由伏隔核和嗅结节组成。[4] [10]伏隔核由伏隔核和伏隔核组成,它们因神经群体而不同。嗅球结节接受来自嗅球的输入,但未被证明在处理气味方面起作用。[10]在非灵长类物种中,包括Calleja岛屿。[11]腹侧纹状体与边缘系统相关,并且被认为是决策和奖励相关行为的重要部分。[12] [13]
背侧纹状体由尾状核和壳核组成。
染色可以将纹状体分化为不同的区域,即纹状体或斑块和周围的基质;这在乙酰胆碱酯酶和钙结合蛋白的组分上尤其明显。对背侧纹状体进行了更多的研究,但在腹侧纹状体中也发现了隔室。在背侧纹状体中,纹状体占纹状体体积的10-15%。[14]
细胞类型

树突状棘刺在纹状体的中型多刺神经元上
纹状体中的细胞类型包括:
中型多刺神经元(MSNs),是纹状体的主要神经元。[2]它们是GABA能的,因此被归类为抑制性神经元。中型多刺投射神经元占人纹状体总神经元群体的95%[2]。中型多刺神经元有两种特征类型:D1型MSN和D2型MSN。[2] [4] [15] MSN亚群包含D1型和D2型受体,大约40%的纹状体MSN表达DRD1和DRD2 mRNA [2] [4] [15]
胆碱能中间神经元释放乙酰胆碱,其在纹状体中具有多种重要作用。在人类,其他灵长类动物和啮齿动物中,这些中间神经元对突出的环境刺激作出反应,其定型反应与黑质多巴胺能神经元的反应在时间上一致。[16] [17]大型阿司匹林胆碱能中间神经元本身受多巴胺通过D5多巴胺受体的影响。[18]
有许多类型的GABAergic中间神经元。[19]最着名的是表达小球蛋白的中间神经元,也称为快速中间神经元,它们参与主要神经元的强大前馈抑制。[20]此外,还有GABAergic中间神经元表达酪氨酸羟化酶,[21]生长抑素,一氧化氮合酶和神经肽-y。最近,已经详细描述了两种表达神经肽y的GABA能中间神经元,[22]其中一种将胆碱能中间神经元的同步活动转化为主要神经元的抑制[23]。纹状体的这些神经元不均匀分布。[19]
大脑中存在两个神经发生区域 - 侧脑室的脑室下区和齿状回。在与纹状体相邻的侧脑室中形成的成神经细胞整合在纹状体中。[24] [25]在缺血性中风后的人纹状体中已经注意到这一点。对纹状体造成的损伤刺激成神经细胞从脑室下区迁移到纹状体,在那里它们分化成成体神经元。[26] SVZ成神经细胞正常通过嗅球,但这种交通在缺血性中风后转移到纹状体。然而,很少有新发现的神经元存活。[27]
输入

额叶皮层到纹状体到丘脑通路的简化图 - 前纹状体回路

基底神经节的主要电路概述。纹状体以蓝色显示。图片显示已叠加的2个冠状切片,包括所涉及的基底神经节结构。 +和 - 箭头处的符号分别表示该途径是兴奋的还是抑制的。绿色箭头指的是兴奋性谷氨酸能通路,红色箭头指的是抑制性GABA能通路,绿松石箭头指的是直接通路上兴奋的多巴胺能通路和间接通路的抑制作用。
[28]就细胞轴突而言,最大的连接来自皮层。新皮质的许多部分支配背侧纹状体。投射到纹状体的皮层锥体神经元位于II-VI层,最密集的投影来自V层。[29]它们主要终止于多刺神经元的树突棘。它们是谷氨酸能的,令人兴奋的纹状体神经元。
纹状体被视为具有自己的内部微电路。[30]腹侧纹状体接受来自大脑皮层和边缘结构的多个区域的直接输入,例如杏仁核,丘脑和海马,以及内嗅皮层和颞下回。[31]它的主要输入是基底神经节系统。此外,中脑边缘通路从腹侧被盖区突出到腹侧纹状体的伏隔核[32]。
另一个众所周知的传入是由黑质致密部神经元引起的黑质纹状体连接。虽然皮质轴突主要在多刺神经元的脊柱头上突触,但黑质轴突突触主要在脊柱轴上。在灵长类动物中,丘脑纹状体传入来自丘脑的中央 - 束旁复合体(参见灵长类动物基底神经节系统)。这种传入是谷氨酸能的。真正的层内神经元的参与更加有限。纹状体还接受来自基底神经节的其他元件的传入,例如丘脑底核(谷氨酸能)或外部苍白球(GABA能)。
目标
更多信息:中型多刺神经元
腹侧纹状体的主要输出突出到腹侧苍白球,然后是丘脑的内侧背核,这是前纹状体回路的一部分。此外,腹侧纹状体突出到苍白球和黑质网状物。其他一些产出包括对延长的杏仁核,下丘脑外侧和脑桥脑核的投射。[33]
来自背侧和腹侧组成部分的纹状体输出主要由中型多刺神经元(MSN)组成,这是一种投射神经元,具有两种主要表型:表达D2样受体的“间接”MSN和表达D1的“直接”MSN。类受体[2] [4]
基底神经节的主要核是纹状体,通过纹状体纤维通路直接投射到苍白球。[34]由于有髓纤维,striato-pallidal路径具有发白的外观。该投射依次包括外部苍白球(GPe),内部苍白球(GPi),黑质致密部(SNc)和黑质网状结构(SNr)。这种投射的神经元受到来自背侧纹状体的GABA能神经突触的抑制。在这些目标中,GPe不会在系统外发送轴突。其他人将轴突送到上丘。另外两个包括丘脑的输出,形成两个独立的通道:一个通过苍白球的内部区域到丘脑的腹侧口腔核,从那里到皮质辅助运动区域,另一个通过黑质到腹侧前部。丘脑的核,从那里到额叶皮质和动眼神经皮质。
功能
腹侧纹状体,特别是伏核,主要介导奖赏,认知,强化和动机显着性,而背侧纹状体主要介导涉及运动功能,某些执行功能(如抑制控制和冲动)和刺激反应的认知。学习; [2] [3] [4] [35] [36]存在一定程度的重叠,因为背侧纹状体也是奖励系统的一个组成部分,与伏隔核一起,调节编码与未来奖励获取相关的新运动程序(例如,条件运动对奖励线索的反应)。[3] [35]
代谢型多巴胺受体存在于多刺神经元和皮质轴突末端上。由激活这些多巴胺受体引发的第二信使级联可以在短期和长期内调节突触前和突触后功能。[37] [38]在人类中,纹状体由与奖赏相关的刺激激活,但也受到厌恶,新颖,[39]意外或强烈刺激以及与此类事件相关的线索的激活。[40] fMRI证据表明,纹状体反应的这些刺激物的共同特性在呈现条件下是显著的。[41] [42]许多其他大脑区域和电路也与奖励相关,例如额叶区域。纹状体的功能图揭示了与广泛分布的大脑皮层区域的相互作用,这些区域对于各种功能是重要的。[43]
纹状体和前额皮质之间的相互作用与行为有关,特别是双系统模型提出的青少年发育。[44]
临床意义
帕金森病
帕金森病导致背侧纹状体(和其他基底神经节)的多巴胺能神经支配丧失和一系列后果。纹状体萎缩还涉及亨廷顿氏病和运动障碍,如舞蹈病,舞蹈病和运动障碍。[45]这些也被描述为基底神经节的电路紊乱。[46]
成瘾
成瘾是一种大脑奖赏系统的紊乱,是由于腹侧纹状体D1型中型多刺神经元中转录因子DeltaFosB(ΔFosB)的过度表达而产生的。 ΔFosB是一种诱导型基因,由于反复过量服用成瘾药物或过度暴露于其他成瘾性刺激物而逐渐在伏隔核中表达[47] [48]。
双相情感障碍
已经观察到PDE10A基因的变体的纹状体表达与一些I型双相障碍患者之间的关联。其他基因的变异,DISC1和GNAS,与双相II障碍有关。[49]
自闭症谱系障碍
自闭症谱系障碍(ASD)的特点是认知缺乏能力和对社会系统的理解不足。这种不灵活的行为起源于前额叶皮质以及纹状体回路中的缺陷。[50]纹状体缺陷似乎特别有助于ASD患者的运动,社交和沟通障碍。在通过翻译因子4E的真核起始过表达而诱导具有ASD样表型的小鼠中,已经表明这些缺陷似乎源于在纹状体中储存和处理信息的能力降低,这导致了困难。从形成新的电机模式,以及脱离现有电机模式。[51]
功能障碍
腹侧纹状体功能障碍可导致多种疾病,最显着的是抑郁症和强迫症。由于其参与奖励途径,腹侧纹状体也涉及在成瘾中起关键作用。已经确定腹侧纹状体通过多巴胺能刺激强烈参与介导药物,特别是兴奋剂的增强作用。[52]
历史
在十七和十八世纪,术语“纹状体”被用来指定半球的许多不同的,深的,皮下的元素。[53] 1941年,Cécile和Oskar Vogt通过为纹状体元素构建的所有元素(参见灵长类动物基底神经节系统)提出术语纹状体来简化命名:尾状核,壳核和眼底纹[54]腹侧部分连接两者一起腹侧向内囊的下部。
比较解剖学家比较脊椎动物之间的皮质下结构这一术语neostriatum是伪造的,因为它被认为是纹状体的系统发育上较新的部分。 该术语仍被某些来源使用,包括医学主题标题。[55]
其他动物
在鸟类中,使用的术语是古纹状体,并且在新的鸟类术语列表(截至2002年)中,新纹状体已经改为nidopallium。[56]
在非灵长类物种中,Calleja岛包括在腹侧纹状体中。[11]
另见
Cortico-basal ganglia-thalamo-cortical loop
List of regions in the human brain
Striatopallidal fibres
参考
"Basal ganglia". BrainInfo. Retrieved 16 August 2015.
Yager LM, Garcia AF, Wunsch AM, Ferguson SM (August 2015). "The ins and outs of the striatum: Role in drug addiction". Neuroscience. 301: 529–541. doi:10.1016/j.neuroscience.2015.06.033. PMC 4523218. PMID 26116518. [The striatum] receives dopaminergic inputs from the ventral tegmental area (VTA) and the substantia nigra (SNr) and glutamatergic inputs from several areas, including the cortex, hippocampus, amygdala, and thalamus (Swanson, 1982; Phillipson and Griffiths, 1985; Finch, 1996; Groenewegen et al., 1999; Britt et al., 2012). These glutamatergic inputs make contact on the heads of dendritic spines of the striatal GABAergic medium spiny projection neurons (MSNs) whereas dopaminergic inputs synapse onto the spine neck, allowing for an important and complex interaction between these two inputs in modulation of MSN activity ... It should also be noted that there is a small population of neurons in the NAc that coexpress both D1 and D2 receptors, though this is largely restricted to the NAc shell (Bertran- Gonzalez et al., 2008). ... Neurons in the NAc core and NAc shell subdivisions also differ functionally. The NAc core is involved in the processing of conditioned stimuli whereas the NAc shell is more important in the processing of unconditioned stimuli; Classically, these two striatal MSN populations are thought to have opposing effects on basal ganglia output. Activation of the dMSNs causes a net excitation of the thalamus resulting in a positive cortical feedback loop; thereby acting as a ‘go’ signal to initiate behavior. Activation of the iMSNs, however, causes a net inhibition of thalamic activity resulting in a negative cortical feedback loop and therefore serves as a ‘brake’ to inhibit behavior ... there is also mounting evidence that iMSNs play a role in motivation and addiction (Lobo and Nestler, 2011; Grueter et al., 2013). ... Together these data suggest that iMSNs normally act to restrain drug-taking behavior and recruitment of these neurons may in fact be protective against the development of compulsive drug use.
Taylor SB, Lewis CR, Olive MF (February 2013). "The neurocircuitry of illicit psychostimulant addiction: acute and chronic effects in humans". Subst. Abuse Rehabil. 4: 29–43. doi:10.2147/SAR.S39684. PMC 3931688. PMID 24648786. The DS (also referred to as the caudate-putamen in primates) is associated with transitions from goal-directed to habitual drug use, due in part to its role in stimulus–response learning.28,46 As described above, the initial rewarding and reinforcing effects of drugs of abuse are mediated by increases in extracellular DA in the NAc shell, and after continued drug use in the NAc core.47,48 After prolonged drug use, drug-associated cues produce increases in extracellular DA levels in the DS and not in the NAc.49 This lends to the notion that a shift in the relative engagement from the ventral to the dorsal striatum underlies the progression from initial, voluntary drug use to habitual and compulsive drug use.28 In addition to DA, recent evidence indicates that glutamatergic transmission in the DS is important for drug-induced adaptations and plasticity within the DS.50
Ferré S, Lluís C, Justinova Z, Quiroz C, Orru M, Navarro G, Canela EI, Franco R, Goldberg SR (June 2010). "Adenosine-cannabinoid receptor interactions. Implications for striatal function". Br. J. Pharmacol. 160 (3): 443–453. doi:10.1111/j.1476-5381.2010.00723.x. PMC 2931547. PMID 20590556. Two classes of MSNs, which are homogeneously distributed in the striatum, can be differentiated by their output connectivity and their expression of dopamine and adenosine receptors and neuropeptides. In the dorsal striatum (mostly represented by the nucleus caudate-putamen), enkephalinergic MSNs connect the striatum with the external globus pallidus and express the peptide enkephalin and a high density of dopamine D2 and adenosine A2A receptors (they also express adenosine A1 receptors), while dynorphinergic MSNs connect the striatum with the substantia nigra (pars compacta and reticulata) and the entopeduncular nucleus (internal globus pallidus) and express the peptides dynorphin and substance P and dopamine D1 and adenosine A1 but not A2A receptors ... These two different phenotypes of MSN are also present in the ventral striatum (mostly represented by the nucleus accumbens and the olfactory tubercle). However, although they are phenotypically equal to their dorsal counterparts, they have some differences in terms of connectivity. First, not only enkephalinergic but also dynorphinergic MSNs project to the ventral counterpart of the external globus pallidus, the ventral pallidum, which, in fact, has characteristics of both the external and internal globus pallidus in its afferent and efferent connectivity. In addition to the ventral pallidum, the internal globus pallidus and the substantia nigra-VTA, the ventral striatum sends projections to the extended amygdala, the lateral hypothalamus and the pedunculopontine tegmental nucleus. ... It is also important to mention that a small percentage of MSNs have a mixed phenotype and express both D1 and D2 receptors (Surmeier et al., 1996).
"striatum | Definition of striatum in English by Oxford Dictionaries". Oxford Dictionaries | English. Retrieved 17 January 2018.
Jones, Jeremy. "Corpus striatum | Radiology Reference Article | Radiopaedia.org". radiopaedia.org. Retrieved 17 January 2018.
"Corpus striatum". BrainInfo. Retrieved 16 August 2015.
Telford, Ryan; Vattoth, Surjith (February 2014). "MR Anatomy of Deep Brain Nuclei with Special Reference to Specific Diseases and Deep Brain Stimulation Localization". The Neuroradiology Journal. 27 (1): 29–43. doi:10.15274/NRJ-2014-10004. PMC 4202840. PMID 24571832.
"Striatum definition and meaning | Collins English Dictionary". www.collinsdictionary.com.
Ubeda-Bañon I, Novejarque A, Mohedano-Moriano A, et al. (2007). "Projections from the posterolateral olfactory amygdala to the ventral striatum: neural basis for reinforcing properties of chemical stimuli". BMC Neurosci. 8: 103. doi:10.1186/1471-2202-8-103. PMC 2216080. PMID 18047654.
"Ventral striatum – NeuroLex". neurolex.org. Retrieved 12 December 2015.
"Ventral Striatum Definition – Medical Dictionary". medicaldictionary.net. Retrieved 18 November 2015.
"Ventral Striatum – Medical Definition". www.medilexicon.com. Retrieved 18 November 2015.
Brimblecombe, K. R.; Cragg, S. J. (2017). "The Striosome and Matrix Compartments of the Striatum: A Path through the Labyrinth from Neurochemistry toward Function". Acs Chemical Neuroscience. 8 (2): 235–242. doi:10.1021/acschemneuro.6b00333. PMID 27977131.
Nishi A, Kuroiwa M, Shuto T (July 2011). "Mechanisms for the modulation of dopamine d(1) receptor signaling in striatal neurons". Front Neuroanat. 5: 43. doi:10.3389/fnana.2011.00043. PMC 3140648. PMID 21811441. Dopamine plays critical roles in the regulation of psychomotor functions in the brain (Bromberg-Martin et al., 2010; Cools, 2011; Gerfen and Surmeier, 2011). The dopamine receptors are a superfamily of heptahelical G protein-coupled receptors, and are grouped into two categories, D1-like (D1, D5) and D2-like (D2, D3, D4) receptors, based on functional properties to stimulate adenylyl cyclase (AC) via Gs/olf and to inhibit AC via Gi/o, respectively ... It has been demonstrated that D1 receptors form the hetero-oligomer with D2 receptors, and that the D1–D2 receptor hetero-oligomer preferentially couples to Gq/PLC signaling (Rashid et al., 2007a,b). The expression of dopamine D1 and D2 receptors are largely segregated in direct and indirect pathway neurons in the dorsal striatum, respectively (Gerfen et al., 1990; Hersch et al., 1995; Heiman et al., 2008). However, some proportion of medium spiny neurons are known to expresses both D1 and D2 receptors (Hersch et al., 1995). Gene expression analysis using single cell RT-PCR technique estimated that 40% of medium spiny neurons express both D1 and D2 receptor mRNA (Surmeier et al., 1996).
Goldberg, JA; Reynolds, JN (December 2011). "Spontaneous firing and evoked pauses in the tonically active cholinergic interneurons of the striatum". Neuroscience. 198: 27–43. doi:10.1016/j.neuroscience.2011.08.067. PMID 21925242.
Morris, G; Arkadir, D; Nevet, A; Vaadia, E; Bergman, H (July 2004). "Coincident but distinct messages of midbrain dopamine and striatal tonically active neurons". Neuron. 43 (1): 133–43. doi:10.1016/j.neuron.2004.06.012. PMID 15233923.
Bergson, C; Mrzljak, L; Smiley, J. F.; Pappy, M; Levenson, R; Goldman-Rakic, P. S. (1995). "Regional, cellular, and subcellular variations in the distribution of D1 and D5 dopamine receptors in primate brain". The Journal of Neuroscience. 15 (12): 7821–36. doi:10.1523/JNEUROSCI.15-12-07821.1995. PMID 8613722.
Tepper JM, Tecuapetla F, Koós T, Ibáñez-Sandoval O. Front Neuroanat. 2010 Dec 29;4:150. doi: 10.3389/fnana.2010.00150. PMID 21228905
Koós, T; Tepper, JM (May 1999). "Inhibitory control of neostriatal projection neurons by GABAergic interneurons". Nat Neurosci. 2 (5): 467–72. doi:10.1038/8138. PMID 10321252.
Ibáñez-Sandoval, O; Tecuapetla, F; Unal, B; Shah, F; Koós, T; Tepper, JM (2010). "Electrophysiological and morphological characteristics and synaptic connectivity of tyrosine hydroxylase-expressing neurons in adult mouse striatum". J Neurosci. 30 (20): 6999–7016. doi:10.1523/JNEUROSCI.5996-09.2010. PMC 4447206. PMID 20484642.
Ibáñez-Sandoval, O; Tecuapetla, F; Unal, B; Shah, F; Koós, T; Tepper, JM (November 2011). "A novel functionally distinct subtype of striatal neuropeptide Y interneuron". J Neurosci. 31 (46): 16757–69. doi:10.1523/JNEUROSCI.2628-11.2011. PMC 3236391. PMID 22090502.
English DF, Ibanez-Sandoval O, Stark E, Tecuapetla F, Buzsáki G, Deisseroth K, Tepper JM, Koos T. Nat Neurosci. 2011 Dec 11;15(1):123-30. doi: 10.1038/nn.2984. PMID 22158514
Ernst, Aurélie; Alkass, Kanar; Bernard, Samuel; Salehpour, Mehran; Perl, Shira; Tisdale, John; Possnert, Göran; Druid, Henrik; Frisén, Jonas (February 2014). "Neurogenesis in the Striatum of the Adult Human Brain". Cell. 156 (5): 1072–1083. doi:10.1016/j.cell.2014.01.044. PMID 24561062.
Inta, D; Lang, U E; Borgwardt, S; Meyer-Lindenberg, A; Gass, P (16 February 2016). "Adult neurogenesis in the human striatum: possible implications for psychiatric disorders". Molecular Psychiatry. 21 (4): 446–447. doi:10.1038/mp.2016.8. PMID 26878892.
Kernie, SG; Parent, JM (February 2010). "Forebrain neurogenesis after focal Ischemic and traumatic brain injury". Neurobiology of Disease. 37 (2): 267–74. doi:10.1016/j.nbd.2009.11.002. PMC 2864918. PMID 19909815.
Yamashita, T; Ninomiya, M; Hernández Acosta, P; García-Verdugo, JM; Sunabori, T; Sakaguchi, M; Adachi, K; Kojima, T; Hirota, Y; Kawase, T; Araki, N; Abe, K; Okano, H; Sawamoto, K (14 June 2006). "Subventricular zone-derived neuroblasts migrate and differentiate into mature neurons in the post-stroke adult striatum". The Journal of Neuroscience. 26 (24): 6627–36. doi:10.1523/jneurosci.0149-06.2006. PMID 16775151.
"ScienceDirect". Brain Research. 175 (2): 191–217. 19 October 1979. doi:10.1016/0006-8993(79)91001-1. PMID 314832. Retrieved 14 April 2019.
Rosell A, Giménez-Amaya JM (1999). "Anatomical re-evaluation of the corticostriatal projections to the caudate nucleus: a retrograde labeling study in the cat". Neurosci Res. 34 (4): 257–69. doi:10.1016/S0168-0102(99)00060-7. PMID 10576548.
Stocco, Andrea; Lebiere, Christian; Anderson, John R. (2010). "Conditional Routing of Information to the Cortex: A Model of the Basal Ganglia's Role in Cognitive Coordination". Psychological Review. 117 (2): 541–74. doi:10.1037/a0019077. PMC 3064519. PMID 20438237.
"Ventral striatum – NeuroLex". neurolex.org. Retrieved 12 December 2015.
"Icahn School of Medicine | Neuroscience Department | Nestler Lab | Brain Reward Pathways". neuroscience.mssm.edu. Retrieved 12 December 2015.
Trevor W. Robbins; Barry J. Everitt (April 1992). "Functions of dopamine in the dorsal and ventral striatum". Seminars in Neuroscience. 4 (2): 119–127. doi:10.1016/1044-5765(92)90010-Y.
Pujol, S.; Cabeen, R.; Sébille, S. B.; Yelnik, J.; François, C.; Fernandez Vidal, S.; Karachi, C.; Zhao, Y.; Cosgrove, G. R.; Jannin, P.; Kikinis, R.; Bardinet, E. (2016). "In vivo Exploration of the Connectivity between the Subthalamic Nucleus and the Globus Pallidus in the Human Brain Using Multi-Fiber Tractography". Frontiers in Neuroanatomy. 10: 119. doi:10.3389/fnana.2016.00119. PMC 5243825. PMID 28154527.
Malenka RC, Nestler EJ, Hyman SE (2009). Sydor A, Brown RY (eds.). Molecular Neuropharmacology: A Foundation for Clinical Neuroscience (2nd ed.). New York: McGraw-Hill Medical. pp. 147–148, 321, 367, 376. ISBN 978-0-07-148127-4. VTA DA neurons play a critical role in motivation, reward-related behavior (Chapter 15), attention, and multiple forms of memory. This organization of the DA system, wide projection from a limited number of cell bodies, permits coordinated responses to potent new rewards. Thus, acting in diverse terminal fields, dopamine confers motivational salience (“wanting”) on the reward itself or associated cues (nucleus accumbens shell region), updates the value placed on different goals in light of this new experience (orbital prefrontal cortex), helps consolidate multiple forms of memory (amygdala and hippocampus), and encodes new motor programs that will facilitate obtaining this reward in the future (nucleus accumbens core region and dorsal striatum). In this example, dopamine modulates the processing of sensorimotor information in diverse neural circuits to maximize the ability of the organism to obtain future rewards. ...
Functional neuroimaging in humans demonstrates activation of the prefrontal cortex and caudate nucleus (part of the striatum) in tasks that demand inhibitory control of behavior. ...
The brain reward circuitry that is targeted by addictive drugs normally mediates the pleasure and strengthening of behaviors associated with natural reinforcers, such as food, water, and sexual contact. Dopamine neurons in the VTA are activated by food and water, and dopamine release in the NAc is stimulated by the presence of natural reinforcers, such as food, water, or a sexual partner. ...
The NAc and VTA are central components of the circuitry underlying reward and memory of reward. As previously mentioned, the activity of dopaminergic neurons in the VTA appears to be linked to reward prediction. The NAc is involved in learning associated with reinforcement and the modulation of motoric responses to stimuli that satisfy internal homeostatic needs. The shell of the NAc appears to be particularly important to initial drug actions within reward circuitry; addictive drugs appear to have a greater effect on dopamine release in the shell than in the core of the NAc.
Kim, BaekSun; Im, Heh-In (2018). "The role of the dorsal striatum in choice impulsivity". Annals of the New York Academy of Sciences. doi:10.1111/nyas.13961. PMID 30277562.
Greengard, P (2001). "The neurobiology of slow synaptic transmission". Science. 294 (5544): 1024–30. doi:10.1126/science.294.5544.1024. PMID 11691979.
Cachope, R; Cheer (2014). "Local control of striatal dopamine release". Frontiers in Behavioral Neuroscience. 8: 188. doi:10.3389/fnbeh.2014.00188. PMC 4033078. PMID 24904339.
UCL (25 June 2008). "Adventure - it's all in the mind, say UCL neuroscientists". UCL News.
Volman, S. F.; Lammel; Margolis; Kim; Richard; Roitman; Lobo (2013). "New insights into the specificity and plasticity of reward and aversion encoding in the mesolimbic system". Journal of Neuroscience. 33 (45): 17569–76. doi:10.1523/JNEUROSCI.3250-13.2013. PMC 3818538. PMID 24198347.
LUNA, BEATRIZ; SWEENEY, JOHN A. (1 June 2004). "The Emergence of Collaborative Brain Function: fMRI Studies of the Development of Response Inhibition". Annals of the New York Academy of Sciences. 1021 (1): 296–309. doi:10.1196/annals.1308.035. PMID 15251900.
"Department of Physiology, Development and Neuroscience: About the Department".
Choi EY, Yeo BT, Buckner RL (2012). "The organization of the human striatum estimated by intrinsic functional connectivity". Journal of Neurophysiology. 108 (8): 2242–2263. doi:10.1152/jn.00270.2012. PMC 3545026. PMID 22832566.
Steinberg, Laurence (April 2010). "A dual systems model of adolescent risk-taking". Developmental Psychobiology. 52 (3): 216–224. doi:10.1002/dev.20445. ISSN 1098-2302. PMID 20213754.
Walker FO (January 2007). "Huntington's disease". Lancet. 369 (9557): 218–28. doi:10.1016/S0140-6736(07)60111-1. PMID 17240289.
Delong, M. R.; Wichmann, T. (2007). "Circuits and circuit disorders of the basal ganglia". Archives of Neurology. 64 (1): 20–4. doi:10.1001/archneur.64.1.20. PMID 17210805.
Nestler EJ (December 2013). "Cellular basis of memory for addiction". Dialogues Clin. Neurosci. 15 (4): 431–443. PMC 3898681. PMID 24459410.
Olsen CM (December 2011). "Natural rewards, neuroplasticity, and non-drug addictions". Neuropharmacology. 61 (7): 1109–22. doi:10.1016/j.neuropharm.2011.03.010. PMC 3139704. PMID 21459101.
Table 1
McDonald, M-L; MacMullen, C; Liu, D J; Leal, S M; Davis, R L (October 2012). "Genetic association of cyclic AMP signaling genes with bipolar disorder". Translational Psychiatry. 2 (10): e169. doi:10.1038/tp.2012.92. PMC 3565822. PMID 23032945.
Fineberg, NA; Potenza, MN (November 2009). "Probing compulsive and impulsive behaviors, from animal models to endophenotypes: a narrative review" (PDF). Neuropsychopharmacology. 35 (3): 591–604. doi:10.1038/npp.2009.185. PMC 3055606. PMID 19940844.
Santini, E; Huynh, TN; MacAskil, A; Carter, A (January 2013). "Exaggerated translation causes synaptic and behavioural aberrations associated with autism". Nature. 493: 411–415. doi:10.1038/nature11782.
Everitt, Barry J.; Robbins, Trevor W. (1 November 2013). "From the ventral to the dorsal striatum: Devolving views of their roles in drug addiction". Neuroscience & Biobehavioral Reviews. Honoring Ann Kelley. 37 (9, Part A): 1946–1954. doi:10.1016/j.neubiorev.2013.02.010. PMID 23438892.
Raymond Vieussens, 1685
"NeuroNames Ancillary: fundus striati". braininfo.rprc.washington.edu. Retrieved 17 January 2018.
Neostriatum at the US National Library of Medicine Medical Subject Headings (MeSH)
"New Terminology for the Neostriatum". www.avianbrain.org. Retrieved 17 January 2018. |